


Genética
As bases bioquímicas da genômica

Texto: Isabela Fonseca, Marta Fonseca Martins, Sabine Wohlres Viana, Marcos Vinícius Gualberto Barbosa da Silva, Marco Antônio Machado
O DNA é a abreviação de ácido desoxirribonucleico e é conhecido como a molécula da vida, responsável pela transmissão de informações genéticas de uma geração para outra. Ele está localizado no núcleo de todas as células de um indivíduo, por isso, para a realização de um exame de DNA basta que qualquer material que contenha alguma célula com núcleo seja coletado, como, por exemplo, bulbos capilares, secreções, sangue ou tecido.
As chamadas bases nitrogenadas (adenina - A, timina - T, citosina - C e guanina - G) fazem parte de sua constituição, por isso o DNA é comumente representado por uma sequência de letras A, T, C e G, como pode ser observado na Figura 1. As bases ficam pareadas para formar a estrutura do DNA (dupla hélice), sendo que A pareia com T e C pareia com G. O conjunto dessas sequências forma o genoma. Foi estimado que o genoma de bovinos contivesse três bilhões de pares de bases (A e T; G e C).

Gene é uma sequência dessas letras capaz de gerar um produto funcional, que pode ser uma proteína ou um RNA (abreviação de ácido ribonucleico). Ele é a unidade que será herdada de uma geração para a outra. O gene candidato, termo comumente visto em anúncios de serviços genéticos e de melhoramento animal, é o gene cuja função foi deduzida por meio de técnicas de genética molecular e que controla uma característica, seja simples como cor de pelagem ou complexa, como produção de leite, resistência a carrapatos ou peso ao nascer. Estima-se que o genoma bovino contenha aproximadamente 22.000 genes.
Cada óvulo contém uma cópia de cada gene da espécie, a mesma coisa acontece com os espermatozoides, por isso, durante a formação de um indivíduo ele receberá duas cópias de cada gene (uma cópia da mãe e outra do pai). As formas alternativas que um gene pode ter são chamadas de alelos. Por exemplo, na espécie humana existe o gene para o grupo sanguíneo (apenas um gene), o qual pode estar na forma de três alelos: A, B e i. Apesar de existirem três alelos, como nós recebemos apenas duas cópias de cada gene (uma cópia da mãe e outra do pai), não é possível um indivíduo normal possuir as três formas alternativas do gene. Portanto, as combinações possíveis são: AA, AB, Ai, BB, Bi e ii. Essas seis combinações possíveis são chamadas de genótipo, o qual é possível ter acesso por meio de técnicas moleculares. Mas, se o genótipo do indivíduo for AA ou Ai, ele terá o sangue do tipo “A” - este é chamado de fenótipo, ou seja, o que é possível de ser observado ou a expressão da característica. Da mesma forma, se o indivíduo for BB ou Bi, ele terá o sangue do tipo “B”; se for AB, terá o sangue do tipo “AB” e, por fim, se o genótipo for ii terá o sangue do tipo “O”.
Em bovinos, a presença ou não de chifres é controlada por um gene, com dois alelos: M, que determina a ausência de chifres e m, que determina a presença deles. O alelo para mocho (M) é dominante sobre o alelo para presença de chifres (m), ou seja, na presença do alelo M, o animal será mocho, mesmo que o outro alelo determine a presença de chifres. Da mesma forma, se um animal tiver os dois alelos para mocho (MM), ele será mocho. No entanto, caso o indivíduo possua os dois alelos para presença de chifres (mm), ele terá chifres, uma vez que, nesse caso, não existe o alelo dominante. Percebe-se que não há como determinar visualmente se um animal com fenótipo mocho possui o genótipo MM ou Mm.
O genótipo normalmente se refere a uma característica em particular (tipo sanguíneo, cor dos olhos, altura, presença de chifres), enquanto o fenótipo é resultado da expressão do conjunto de genes de um organismo, modulada pelos fatores ambientais. Como o fenótipo é uma combinação entre genótipo e ambiente, organismos que tenham um mesmo genótipo não necessariamente terão um fenótipo igual. Do mesmo modo, organismos com mesmo fenótipo não têm o mesmo genótipo, obrigatoriamente.
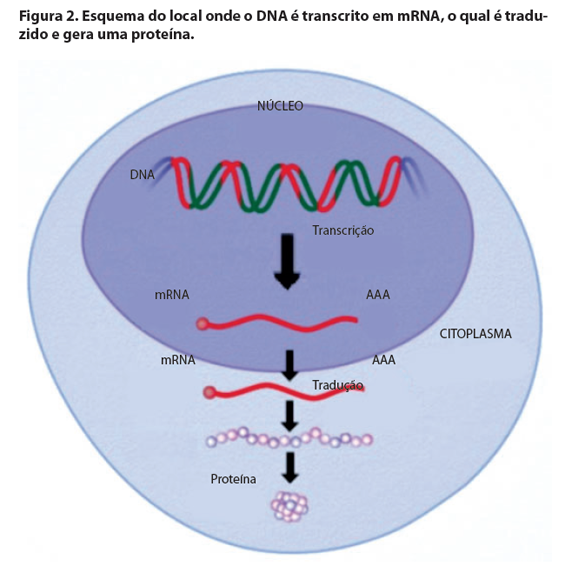
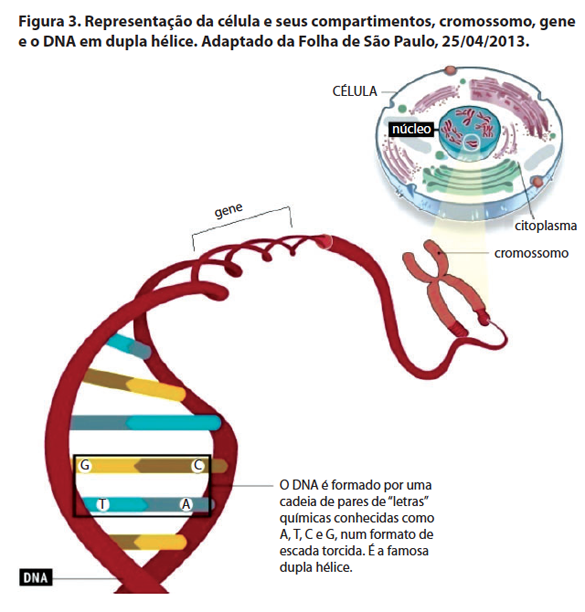
Para que as informações contidas no DNA sejam traduzidas em proteínas e gerem o fenótipo, é preciso que o DNA seja transcrito em RNA (ácido ribonucleico). Isso porque o DNA localiza-se no núcleo da célula, no entanto, a proteína é sintetizada no citoplasma. O RNA mensageiro (mRNA) é transcrito no núcleo, usando como molde o DNA e vai para o citoplasma para ser traduzido em proteína, produto final de um gene (Figura 2). O cromossomo nada mais é do que o DNA condensado (Figura 3).
Entre os animais de um determinado rebanho, existem pequenas alterações naturais na sequência do DNA, sendo que cada animal possui um padrão específico destas variações. Estas alterações naturais funcionam como marcadores moleculares dos alelos, os quais podem possuir efeitos positivos ou negativos numa determinada característica de interesse. A seleção de animais, que possuem alelos favoráveis para os genes, controlando as características em questão, baseada na avaliação direta de seu DNA, é denominada de Seleção Assistida por Marcadores (SAM).
Um marcador molecular é uma sequência de DNA ou um tipo de proteína que está associada a uma parte do genoma. Os melhores marcadores moleculares são os baseados em DNA, uma vez que estas sequências são altamente específicas entre os indivíduos. Um marcador de DNA pode ser usado como ferramenta para a seleção de animais mais produtivos e resistentes às doenças, por exemplo o marcador molecular para a doença Deficiência de Adesão Leucocitária Bovina (BLAD - do inglês Bovine Leukocyte Adhesion Deficiency). Uma única mutação no gene CD18 em que houve a troca de um A por G causou esta doença hereditária comum na raça Holandesa. Animais homozigotos para esta mutação, ou seja, que receberam os dois alelos (um da mãe e outro do pai) em que houve a troca de A por G num determinado local do gene CD18 (Figura 4), apresentam crescimento retardado, perda de dentes, comprometimento do sistema imune e morrem ainda novos, geralmente, de pneumonia. Animais heterozigotos (possuem apenas um dos alelos em que houve a troca) apresentam desenvolvimento normal e podem transmitir a mutação para a progênie. Caso animais portadores cruzem entre si a probabilidade de um animal ser afetado pela doença é 25%. Por isso, orienta-se que animais portadores do alelo BLAD devem ser acasalados com outros não portadores. No entanto, para saber se um animal normal é portador ou não de um alelo com a mutação, é necessária a realização de um exame de DNA, neste caso a genotipagem do gene CD18.

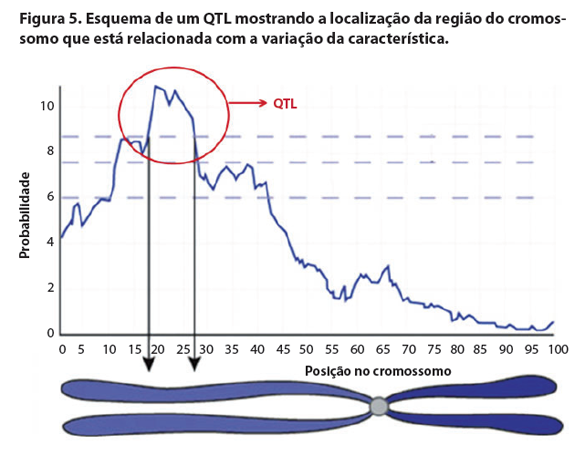
Como pode ser observado, esta doença é causada por uma mutação em apenas um gene e só há dois fenótipos possíveis: normal ou doente. Este tipo de característica é chamado na genética como “característica qualitativa”, a qual é regulada por um ou poucos genes e praticamente independem do ambiente para se manifestar. Porém, a maioria das características observadas é complexa, regulada por vários genes, sofre muita influência do ambiente e exibe variações contínuas (altura, cor da pelagem, resistência a doenças, produção de leite, peso ao desmame, etc.). Por isso, é necessário outro tipo de marcador para estes caracteres quantitativos, chamado QTL (Quantitative Trait Loci). QTL são as regiões do DNA que estão relacionadas com a variação fenotípica das características quantitativas. Neste caso, não existe apenas um gene responsável pela variação da característica, mas sim vários genes, ou uma região do genoma que responde em maior parte por esta variação. Por exemplo, para resistência à mastite, existem trabalhos científicos que localizaram QTLs em praticamente todos os cromossomos de bovinos, sendo que alguns QTLs apresentam maiores efeitos sobre a característica em relação a outros.
Os dados moleculares oriundos destes marcadores podem ser associados aos dados fenotípicos e utilizados na SAM. Este tipo de seleção apresenta inúmeras vantagens quando comparada ao melhoramento convencional, por exemplo: aumento do ganho genético por geração por aumentar a precisão da seleção; não é limitada pelo sexo, logo os animais machos também podem ser selecionados para produção de leite; e permite a seleção de animais jovens. No entanto, mesmo com as vantagens que a SAM possibilita, é interessante considerar também as informações obtidas com as metodologias convencionais utilizadas nos programas de melhoramento das raças, visando maximizar o ganho genético.
Até recentemente, o número de marcadores de DNA identificados no genoma de bovinos era limitado e o custo de genotipagem era alto. Porém, com o advento dos marcadores tipo SNP (polimorfismos de base única, do inglês Single Nucleotide Polymorphism), a seleção será feita com o auxílio de milhares de marcadores distribuídos por todo o genoma, a chamada “Seleção Genômica”. O SNP, marcador mais utilizado atualmente, é resultante da alteração de um único par de bases em uma determinada posição. Como este marcador é muito abundante no genoma, é possível automatizar o processo e, com isso, diminuir o custo de genotipagem por animal. A seleção genômica usa associações de grande número de marcadores SNPs espalhados pelo genoma com as características fenotípicas, como os dados de produção, por exemplo. Com base nestas associações, é possível estimar o desempenho da progênie de um reprodutor de acordo com as informações contidas no seu DNA.
No passado, para escolher entre dois reprodutores jovens para entrar no Teste de Progênie, filhos da mesma vaca e do mesmo touro, a única informação disponível era a informação de pedigree, e não havia modo para determinar se o desempenho de um dos animais seria melhor ou pior do que a média dos seus pais. Assim, seriam necessários cerca de sete anos para que, com base no desempenho de suas progênies, esses animais fossem avaliados. Atualmente, em virtude da relação existente entre os marcadores moleculares e os genes relacionados às características de interesse econômico, é possível estimar com segurança o provável desempenho de um animal logo após o seu nascimento. Além disso, a avaliação genômica também considera informações de pedigree e pode incluir ainda informações da progênie. Ou seja, as provas genômicas baseiam-se nos mesmos princípios que as provas tradicionais, mas adicionam as informações do DNA para incrementar a confiabilidade e redução do tempo de prova.
No Brasil, um projeto em andamento na Embrapa Gado de Leite visa o desenvolvimento de estratégias para a implementação da seleção genômica para as raças Gir, Guzerá e Girolando. O objetivo é entregar aos criadores e centrais de inseminação artificial as ferramentas e o conhecimento necessários para facilitar o uso da seleção genômica para imediata aplicação nos Testes de Progênie das raças.
Os recentes avanços no campo da biotecnologia permitiram a incorporação das informações de marcadores moleculares nos programas de seleção e acasalamento em Gado de Leite. O conhecimento das informações sobre o genótipo de animais tem grande importância estratégia e valor econômico, pois permite identificar os animais de maior potencial de produção de leite, gordura e proteína e portadores de alelos para doenças hereditárias. De posse dessas informações, o produtor pode orientar acasalamentos, a escolha de sêmen e a aplicação da seleção assistida por marcadores moleculares para a melhoria genética do seu rebanho.










Enviar comentário